Many natural processes consist of networks of
interacting elements which affect each other's state over time, the
dynamics depending on the pattern of connections and the updating
rules for each element. Genomic regulatory networks are arguably
networks of this sort. An attempt to understand genomic networks
would benefit from the context of a general theory of discrete
dynamical networks which is currently emerging. A key notion here is
global dynamics, whereby state-space is organized into basins of
attraction, objects that have only recently become accessible by
computer simulation of idealized models[12, 13],
in particular ``random Boolean networks''. Cell types have been
explained as attractors in genomic networks[5], where
the network architecture is biased to achieve a balance between
stability and adaptability in response to perturbation[3].
Based on computer simulations using the software
Discrete Dynamics
Lab (DDLab)
[15], these ideas are described, as well as
order-chaos measures on typical trajectories that
further characterize network dynamics.
Processes consisting of concurrent networks of interacting elements
which affect each other's state over time occur in a wide variety of
natural systems, the dynamics depending both on the pattern of
connections (wiring) and on the update rules for each element. The
behavior of such complex feedback webs is difficult to treat
analytically by classical mathematics using its tools of partial
differential equations and continuous dynamics. Instead, a theoretical
understanding of these networks depends on numerical simulations of
idealized computer models known as discrete dynamical networks.
An example are cellular automata, a powerful yet simple class of
network, characterized by a universal rule and uniform nearest
neighbor wiring, which are used to study processes in physical
systems such as reaction-diffusion[11], and
self-organization by the emergence of coherent interacting
structures[14]. By contrast, biological systems such as
neural, immune or genomic networks require models where wiring and
rules are unconstrained. These more general ``random Boolean
networks'' (RBN) were proposed as models of genomic regulatory
networks by Kauffman[5, 6] and have roots in
Ashby's work on dynamical systems and cybernetics[1].
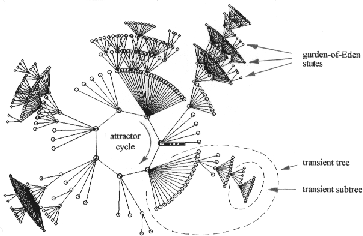
Figure 1:
A basin of attraction (one of 15) of a random Boolean network
(n=13, k=3) defined in figure 4. The basin
links 604 states, of which 523 are garden-of-Eden states. The
attractor has period 7, and one of the attractor states is
shown as a bit pattern. The direction of time is inwards from
garden-of-Eden states to the attractor, then clock-wise.
A key notion underlying the behavior of discrete dynamical networks
is that they organize their state-space into a set of basins of
attraction, object which sum up the network's global dynamics. The
terminology is borrowed from classical continuous dynamical systems.
Basins of attraction can be investigated by running networks forward
from many initial states to discover the range of attractors present.
More recently, exact representations of basins of attraction have
become accessible, where new algorithms allow the network to be run
``backwards'' to disclose all possible historical
paths[12, 13, 16].
The cells of living organisms differentiate within the developing
embryo into the various cell types that form tissues by a process that
is regulated at the molecular level by DNA sequences, encoding genes
that produce proteins that regulate other genes. All eukaryotic cells
in an organism carry an identical set of genes, some of which are
expressed others not. A cell type is defined by the particular subset
of genes that are expressed. The gene expression pattern of a cell
needs to be stable but also adaptable. What then is the mechanism that
maintains the several hundred alternative stable patterns of gene
expression of the various cell types making an organism?
Genes regulate each other's activity by coding for transcription
factors, which may enhance or repress the expression of other genes by
binding (possibly in combination) at particular sites. Though a
particular gene directly regulates just a small set of other genes,
those genes regulate other genes in turn, so a gene will indirectly
influence the activity of many genes downstream. Conversely, a
particular gene is indirectly influenced by many genes upstream. A
gene may directly or indirectly contribute to regulating itself. The
result is a genomic regulatory network, a complex feedback web of
genes turning each other on and off. This may be interpreted as an
idealized dynamical system of model genes with directional links or
``wires'' (transcription factors), updating their (on-off) state in
parallel, according to the combinatorial logic of their
inputs[8], Kauffman's random Boolean
networks[5, 6].
There is justified debate as to whether parallel (synchronous)
updating, and the on-off characterization of genes, are valid
idealizations when applied to real genomic networks, given that
transcription is asynchronous and driven at different rates. However,
gene activity at the molecular scale consists of discrete events
occurring concurrently. Variable protein concentrations can be
accounted for by genes being on for some fraction of a given time
span. The RBN idealization is arguably a valid starting point for
gaining insights into gene network dynamics.
When genomic regulatory networks are seen from a discrete dynamical
network perspective, the problem of understanding how the range of
stable cell types can exist with identical genes becomes more
understandable. Cell types can be defined as the separate attractors
or basins of attraction into which network dynamics settles from
various initial states. Trajectories leading to attractors can be
seen as the pathways of differentiation.
In a cell type's gene expression pattern over a span of time (i.e. its
space-time pattern), a particular gene may, broadly speaking, be
either on, off, or changing. If a large proportion of the genes are
changing, chaotic dynamics, the cell will be unstable. On the other
hand, dynamics that settles to a pattern where a large proportion of
the genes are permanently on or off (frozen) may be too inflexible for
adaptive behavior. Cells constantly need to adapt their gene
expression pattern in response to a variety of hormone and
growth/differentiation factors from nearby cells. The definition of a
cell type may be more correctly expressed as a set of closely related
gene expression patterns, allowing an essential measure of flexibility
in behavior. Too much flexibility might allow a perturbation to flip
the dynamics into a different basin of attraction, a different cell
type such as a cancer cell, or a bone cell to a fat cell. The
conjecture is that the appropriate dynamical regime has evolved to
find a delicate balance between stable on/off regions and dynamically
changing regions. This can be visualized as a balance between order
and chaos with its particular notion of a phase transition.
Figure 2: RBN architecture. Each element in the network synchronously
updates its value according to the values in a pseudo-neighborhood,
set by single wire couplings to arbitrarily located network elements
at the previous time-step. Each network element may have a different
wiring, rule and number of input wires k. The system is iterated.
Each element (or model gene) in a RBN may have a different wiring
and/or rule scheme (but not necessarily), and also a different number,
k, of input ``wires'' from other network elements. Most work on RBN
has assumed networks with uniform k, but recent investigations of
genomic regulatory networks require setting up networks with
particular proportions of different k inputs to match data from cell
biology[3]. Mixed k networks are also required to check
solutions to the inverse problem, where the network architecture is
deduced from some degree of knowledge of the basin of attraction
field[9].
Figure 3: RBN 1d
space-time pattern, n=150, k=5. Time proceeds from the top
down. Wiring and rules were initially set at random resulting
in chaotic dynamics. Middway in the space-time pattern the
rules were adjusted to set canalizing inputs at 52% resulting
in the rapid emergence of frozen elements, a sign of order.
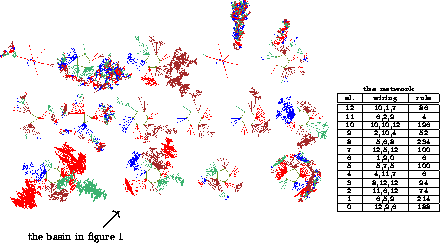
Figure 4:
The basin of attraction field of a random Boolean network (n=13,
k=3). The states in state space are organized into
15 basins, with attractor periods ranging between 1 and 7. The
number of states in each basin is: 68, 984, 784, 1300, 264, 76, 316,
120, 64, 120, 256, 2724, 604, 84, 428. Figure 1 shows
the arrowed basin in more detail. Right: the network's wiring/rule
scheme.
RBN imply a value range of 2, where each element has a value of
either 0 or 1 (an inactive or active gene), but the attractor basin
approach would apply equally to networks with more than two values.
The state of a network of n elements is the pattern resulting from
the values assigned to each element, from a finite range of values v
(usually v=2). Each element synchronously updates its value in
discrete time steps. The value of each element at time t+1
depends on its particular Boolean function , applied to a
notional or pseudo-neighborhood, size k, whose values depend on the
wiring, as illustrated in figure 2. The
resulting sequence of states, the network's trajectory, can be shown
as a space-time pattern in one (or two) dimensions as in figure
3.
A pseudo-neighborhood of size k has permutations of values.
The most general expression of the Boolean function or rule is a
lookup table (the rule table) with entries, giving
possible rules. Sub-categories of rules can also be expressed as
simple algorithms, concise AND/OR/NOT logical statements, totalistic
rules[11] or threshold functions. The number of effectively
different rules is reduced by symmetries in the rule
table[12]. By convention the rule table is arranged in
descending order of the values of neighborhoods, and the resulting
bit string converts to the decimal or hexadecimal rule number.
An example of a k=3 rule table for rule 30 (hex 1e),
The system's parameters are set by specifying the wiring and rule for
each of the n network elements. Once set, the networks wiring/rule
scheme is usually fixed over time.
The i-th cell has wiring connections
. Connections are assigned to any
of the n elements in the network, including itself.
Duplicate connections are allowed, giving possible alternative
wiring options. The rule for element i is , assigned from the
alternatives in rule-space. The time evolution of the
i-th element is given by,
.
The number of all possible alternative wiring/rule schemes that can be
assigned to a given RBN of size n and homogeneous connectivity k
is given by[13], .
This may be compared with the number of possible basin of attraction
fields[16], , for a network of size n, which
equals the number of alternative mappings, , for a
set Q of size n, so .
As for a fully wired network where k=n, which can
achieve any mapping, there must be considerable redundancy in
wiring/rule schemes so that many equivalent schemes generate identical
basin of attraction fields. This is indeed the case. For example, if
the pseudo-neighborhood of a network element is re-ordered, there is
an appropriately re-ordered rule table that gives identical behavior,
thus k! equivalent wiring-rule schemes. Myers[7] has
derived expressions for the number of effectively distinct maps, thus
basin of attraction fields for RBN.
The software DDLab[15] allows a network size n
(arranged in one or two dimensions) to be set up with any arrangement
of wiring, rules, and k values. Wiring and rule parameters can be
restricted or biased in various ways. For example wiring can be
confined within a local neighborhood, and the rules can be tuned to a
given fraction of canalising inputs[3, 16] (see
section 10).
The idea of attractor basins in discrete dynamical networks is
summarized in figure 5.
Given an invariant network architecture and the absence of noise, a
discrete dynamical network is deterministic, and follows a unique
trajectory (a sequence of network states) from any initial state. This
non-random walk through a finite state-space must reach an attractor.
When a state that has occurred previously is encountered, the dynamics
become trapped in a perpetual cycle of repetitions which defines the
attractor (state cycle) and its period (minimum one, a stable point),
whereas transients are sequences outside the attractor but leading to
it.
These systems are dissipative. A state may have multiple predecessors
(including none) but just one successor resulting in a convergent
tree-like topology of the flow in state-space. The set of transient
trees rooted on each attractor state defines a basin of attraction
with a topology of trees rooted on cycles (figure
1). Network architecture thus organizes state-space into
a set, or field, of basins of attraction, representing the systems
global dynamics (figure 4).
The capability of constructing attractor basins depends on reverse
algorithms for directly computing the predecessors (also known as
pre-images) of network states[12, 13, 16].
This allows the network's dynamics, in effect, to be run backwards in
time. Backward trajectories will, as a rule, diverge. These
algorithms open up a window on the precise structure of the basins of
attraction of discrete dynamical networks.
Different reverse algorithms apply to networks with different
qualities of connectivity. The most computationally efficient
algorithm applies to 1d networks with cellular automata type
connections, although the rules for each element may differ. An
alternative algorithm is required for RBN with their non-local
connections and possibly mixed k. This algorithm also applies to
cellular automata, which are just a sub-class of RBN.
These methods are in general orders of magnitude faster than the brute
force method, constructing an exhaustive map resulting from network
dynamics. The exhaustive method rapidly becomes computationally
intractable with increasing network size so is limited to very small
systems, but applies to all network types and also allows the
attractor basins of random maps to be constructed[16].
The three independent methods together form a valuable reality check
on the correctness of the computed pre-images and attractor basins.
These methods are applied in DDLab to compute and portray attractor
basins. Previous investigations of attractor basins relied only on
statistical methods. These are also implemented in DDLab and are
appropriate for large networks (see section 10).
To construct a basin of attraction containing a particular state or
``seed'', the network is iterated forward from the seed state until a
repeat is encountered in the trajectory. This identifies the attractor
cycle as the sequence of states from the state that was repeated.
Once the attractor cycle is known (and drawn as a circle of nodes),
the transient tree (if it exists) rooted on each attractor state is
constructed in turn. Using one of the reverse algorithms, the
pre-images of the attractor state are computed, but the pre-image
lying on the attractor itself is deliberately excluded to prevent
endlessly tracing pre-images backwards around the attractor cycle. The
reverse algorithm is then re-applied repeatedly, to the pre-images of
pre-images, until all the ``leaves'' of the transient tree have been
reached. These are states without pre-images, known in automaton
terminology as ``garden-of-Eden'' states. In this way the transient
tree is completely specified.
In a similar way, just a subtree or a fragment of a subtree may be
constructed rooted on a state. However, because a state chosen at
random is very likely to be a garden-of-Eden state, to construct a
subtree it is usually necessary to run the network forward by at least
one step from the initial random state and use the state reached as
the subtree root. Running forward by more steps will reach a state
deeper in the subtree so allow a larger subtree to be constructed.
Attractor basins are portrayed as directed graphs as presented
in[12]. Global states are represented by nodes, by a bit
pattern in 1d or 2d, or as the decimal or hex value of the state. The
nodes are linked by directed arcs. Each node will have zero or more
incoming arcs from its pre-image nodes (in-degree), but because the
system is deterministic, exactly one outgoing arc (one out-degree).
In the graphic convention for drawing attractor
basins[12], the length of transition arcs decreases with
distance away from the attractor, and the diameter of the attractor
cycle approaches an upper limit with increasing period. The forward
direction of transitions is inward from garden-of-Eden states to the
attractor, which is the only closed loop in the basin, and then
clockwise around the attractor cycle (figure 1 shows an
example). Typically, the vast majority of states in a basin of
attraction lie on transient trees outside the attractor, and the vast
majority of these states are garden-of-Eden states.
A given network defines its basin of attraction field, from which
characteristic measures may be taken, such as the number of
attractors, attractor periods, size of basins, characteristic length
of transients and characteristic branching within trees (the density
of leaves, in-degree frequency). These are also measures of
convergence indicating the degree of order-chaos in the dynamics,
where high leaf density, high branching, short transients, and small
attractor cycles indicate order, and the converse indicates chaos.
Figure 6: The distribution of a set of similar states differing by one
bit from the reference state indicated, are highlighted in basins of
attraction. The network is the same as in figures 1 and 2. Only the
basins which contain the set of states and the reference state are
shown, i.e. basins 2, 11, 12 and 13 in figure 2. Following a one bit
perturbation of the reference state, the dynamics may stay in the
same basin or flip to one of the other basins with a predictable
probability.
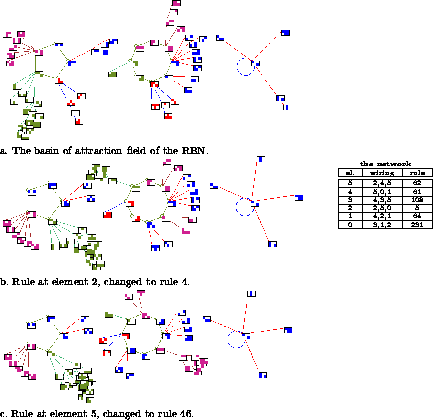
Figure 7: The consequences of mutating a single bit in the rule
table of just one element of a small RBN, n=6, k=3. The state at
each node is shown as a 32 bit pattern.
Above: Network architecture.
The precise structure of attractor basins is of interest as it
reflects the stability of cell types to perturbation, both to the
current state of the network and to the network parameters themselves.
A set of similar states can be specified, for example that differ by
one bit from a reference state (a Hamming distance of one). The
distribution of the set across the basin of attraction field indicates
the network's response to a one bit perturbation to its current state of
activation. The dynamics might remain in the same basin or flip to a
different basin; remain in the same sub-tree or flip to a different
sub-tree. Figure 6
gives an example. An analogous statistical measure is the distribution
of ``damage spread'' resulting from many single bit flips in a large
network[16].
The effect of perturbation to network architecture, mutating the
wiring and/or rules, can be assessed by the affect on the basin of
attraction field, analogous to the role of genotype and phenotype.
Figure 7 shows the
consequences of mutating a single bit in the rule table of just one
element of a small RBN, n=6, k=3. The basin of attraction field is
affected to varying degrees. The example networks are small so that
the pattern at each node can be shown. Larger networks are affected in
analogous ways, although in general the consequences of a one bit
mutation become relatively smaller with increasing network size. However a
particular one bit mutation may cause drastic consequences whatever
the size, such as breaking an attractor cycle. Note that the
consequences of moving a connection wire is usually greater than a one
bit mutation.
Attractors classify state-space into broad categories, the network's
``content addressable'' memory in the sense of
Hopfield[4]. Furthermore, state-space is categorized
along transients, by the root of each subtree forming a hierarchy of
sub-categories. This notion of memory far from the equilibrium
condition of attractors greatly extends the classical concept of
memory by attractors alone[13]. Although such a
discussion would seem only to relate to the area of neural networks,
it also applies to genomic regulatory networks in the sense that they
must ``remember'' and return to their cell types (and sub-types) as they
are continually perturbed by internal and external factors.
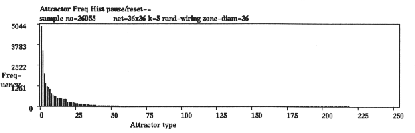
Figure 8: A histogram of attractor types against the frequency of
each type sorted by frequency for a 2d RBN 3636, k=5, with
fully random wiring and a fraction of canalising inputs C=52%.
The data was assembled by running the network forward from a sample
of about 26,000 random initial states and recording the fraction of
different attractors reached, about 220 types in this sample. The
frequency of arriving at each basin of attraction indicates its
relative size, the number of states it contains.
It can be argued that in biological networks such as neural networks
in the brain or networks of genes regulating the differentiation and
adaptive behavior of cells, the topology of attractor basins, the
network's memory, must be just right for effective categorization.
The dynamics need to be sufficiently versatile for adaptive behavior
but short of chaotic to ensure reliable behavior, and this in turn
implies a balance between order and chaos in the network.
Time and memory constraints limit the size of networks when computing
basin of attraction fields, with a practical upper limit of about
n=26, though DDLab allows given sufficient
memory. Single basins of attraction and sub-trees can be computed for
much larger networks. However, data on the basin of attraction field
can be assembled for large networks by statistics on forward dynamics
where the demands on memory are much smaller.
The number and relative size of the different basins in the basin of
attraction field can be inferred from a histogram of the frequency of
reaching different attractor types from many random initial states.
Other data such as attractor length and average run-in length are also
available. In the example in figure 8 the rules scheme is randomly biased to give a fraction of
canalising[3] inputs of 52%. Canalization occurs when a
particular input (0 or 1) on a connection determines a gene's value
irrespective of its other inputs. That connection is then said to be
canalising. This tunes the network to achieve a balance between order
and chaos according to various dynamical measures (see below).
The wiring/rule architecture of RBN can be biased to produce the
percolation of ``frozen'' genes, that have become fixed on or off over
time[6]. These biases have been correlated with data on
biological genomic networks[3] and are understood to
confer stability to the cell type, while the remaining unstable
genes provide adaptability. The dynamics is seen as being marginally
on the ordered side of an order-chaos phase transition in the space of
all possible network architectures according to a number of measures.
In particular, increasing the degree of canalisation of network rules
moves behavior across the transition, though the characteristics of
network wiring also plays a role. Order-chaos
measures[16] listed below may be studied with DDLab,
based on statistics on large networks as well as exact data on small
networks.
Measures on the topology of attractor basins and subtrees such
as their in-degree distribution.
The Derrida plot[2], analogous to the Liaponov
exponent in continuous dynamics, measures the divergence/convergence
of trajectories from pairs of initial states separated by a varying
Hamming distance.
The distribution of ``damage spread'', where the difference
between trajectories from pairs of initial states differing by one
bit is monitored, and the distribution of this damage size is
measured.
The percolation of frozen regions, where the fraction of genes
in a network that have stabilized on or off, or that continue to
flicker is identified. This allows the distribution of ``frozen
skeletons'' to be measured, characteristic frozen patterns that are
treated as quasi-attractors.
The distribution of attractor types as shown in figure 8.
Input-entropy over time relating to single genes, which measures
the frequency with which neighborhoods in the gene's rule-table are
activated over a moving window of time-steps, and the entropy
calculated.
Attractor basins of discrete dynamical networks provide key insights
into a network's global dynamics. Idealizations of genomic regulatory
networks are modeled as random Boolean networks, where the range of
biases in network architecture can be tested against theories and data
from cell biology. A broad range of characteristics, parameters and
measures of global dynamics, as well as dynamics along particular
trajectories, are now open to investigation, including the number of
attractors (cell types), their size distribution and topology,
stability and adaptability to perturbations of both patterns of
activation and network architecture, issues of the network's memory
and the inverse problem[9], and order-chaos measures on
network dynamics. The software Discrete Dynamics Lab[15]
is a tool for these investigations. DDLab continues to be developed
with new capabilities in response to current research ideas.
Harris,E.S., B.K.Sawhill, A.Wuensche, and S.Kauffman,
(1997) ``Biased Eukaryotic Gene Regulation Rules Suggest Genome
Behaviour is Near Edge of Chaos'', Santa Fe Institute Working Paper
97-05-039.
Hopfield,J.J. (1982) ``Neural
networks and physical systems with emergent collective computational
abilities'', Proceedings of the National Academy of Sciences
79:2554-2558.
Somogyi,R., and C.Sniegoski, (1996) ``Modeling the
Complexity of Genetic Networks: Understanding Multigenetic and
Pleiotropic Regulation'', Complexity, Vol.1/No.6,45-63.
Somogyi,R, S.Fuhrman, M.Askenazi, A.Wuensche.,
(1997) ``The Gene Expression Matrix'', in Proceedings of the World
Congress of Non-Linear Analysis, in press.
Wuensche,A., (1994) ``The Ghost in the
Machine'', in Artificial Life III, ed C.G.Langton, Santa Fe
Institute Studies in the Sciences of Complexity, Addison-Wesley.