Homeostatic adaptation
|
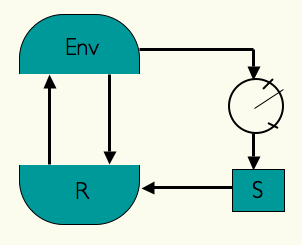
In the 1950's British cybernetician W. Ross Ashby introduced the concept of an ultrastable system in his book Design for a Brain. If an organism needs to keep certain essential variables within bounds (e.g., blood temperature, pH) then a system that triggered internal parametrical changes whenever such variables approach or cross a boundary of viability would, in principle, have a chance of finding new parameters that make the challenged variables return to their viability zones. In the scheme on the left, the organism is represented in interaction with the environment: R represents the behaviour-generating sub-system of the organism which is in a closed
sensorimotor loop with the environment (Env), S represents those parameters that affect R,
and the meter represents the organism's essential variables which can be directly affected by
the environment and in turn affect S by introducing random step-changes in the parameter
values. If these changes produce a new dynamics capable of restoring the essential variables to
their equilibrium through the newly acquired behaviours, the organism will have adapted to the
environmental challenge.
|
|
|
This principle can be applied to adaptation in evolved robots controlled by plastic neural networks.
The artificial evolution of homeostatic controllers for robots was originally inspired by the work of James Garden Taylor (1962) who presented a theory based on
Ashby's concept of ultrastability to account for the fascinating results obtained in experiments on adaptation to distortions of the visual field. Taylor worked in the 1950s at Innsbruck with Erismann and Kohler on adaptation to different sorts of prismatic perturbations including full up/down and left/right inversion. These experiments involved sub jects wearing distorting goggles for a few weeks. They
follow in style similar experiments by Stratton in the 1890s and Ewert in the 1930s. Participants, after several days, experienced behavioural adaptation to those behaviours that they enacted frequently (walking on busy roads, riding a bicycle, and even skiing). In some cases fragmented aspects of perceptual adaptation were reported (world did not look upside down anymore). Upon removal of the goggles a period of re-adaptation was always necessary
|
|
|
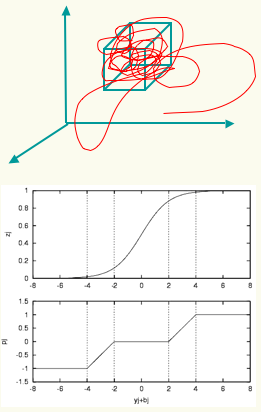
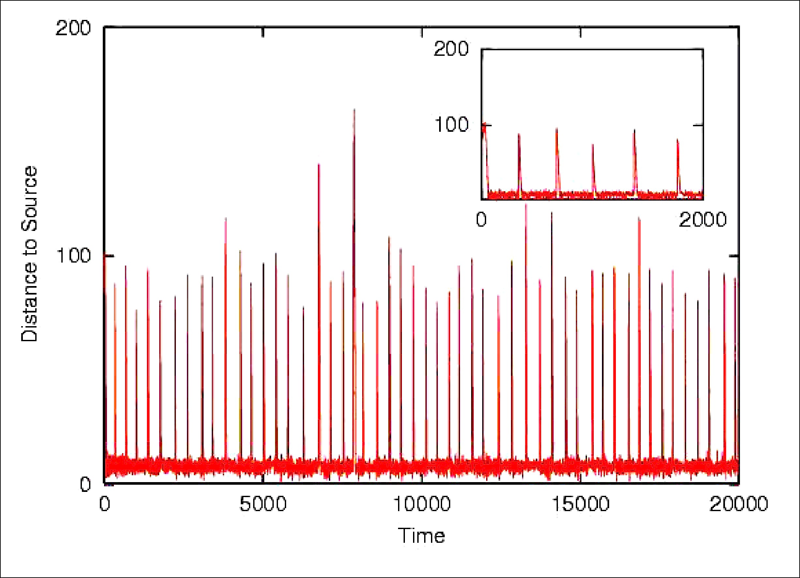
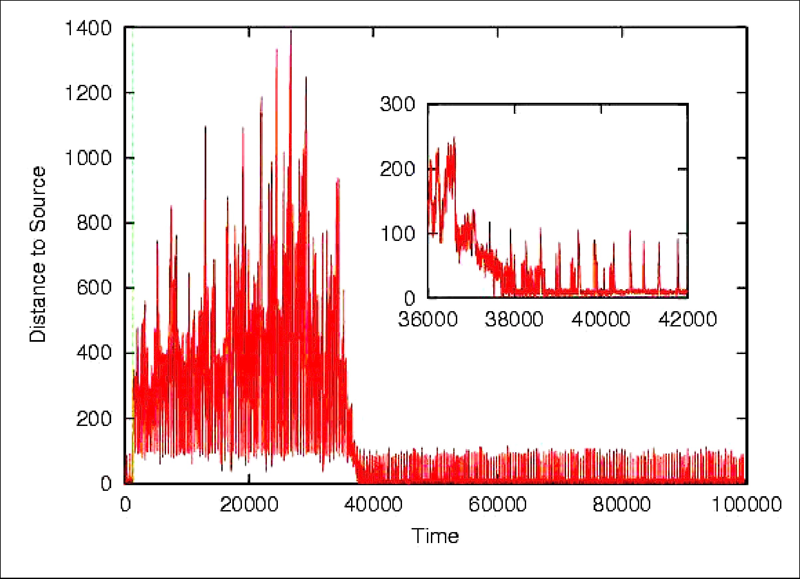
Each neuron in the neurocontroller behaves homeostatically by triggering local plastic changes on incoming synapses whenever it is firing either too much or too little. A viability zone is thus defined in the space of neural activations (left). A mobile agent is evolved to produce a certain behaviour (phototaxis) and to remain as homeostatic as possible. This double fitness requirement (one behavioural and one corresponding to neural stability) generates agents capable adapting to unseen radical sensorimotor perturbations, including the left-right inversion of the visual field (by swapping the position of its sensors). Upon repeated presentations of light sources the agents approaches each new one in turn (below left). If the sensor are swapped (below right) the agents first moves away from the light (as expected). But this induces unstable firing in the neurons which in turn trigger local plastic changes. At some point the network finds its way into the viability zone resulting the the recovery of phototactic behaviour.
The result has interesting implications for understanding the dynamics of adaptive systems capable of long-term interactions with their environment. The homeostatic mechanism produces qualitatively natural developmental patterns such as the presence of critical periods of higher plasticity early in life.
|
|
|
|
Di Paolo, E. A., (2000). Homeostatic
adaptation to inversion of the visual field and other sensorimotor disruptions. From Animals to Animals, Proc. of the Sixth International Conference on the Simulation of Adaptive Behavior, SAB'2000, Paris, J-A. Meyer, A. Berthoz, D. Floreano, H. Roitblat and S W. Wilson (eds), MIT Press, pp. 440 - 449.
Di Paolo, E. A., (2003). Organismically-inspired
robotics: Homeostatic adaptation and natural teleology beyond the closed
sensorimotor loop, in: K. Murase & T. Asakura (Eds) Dynamical
Systems Approach to Embodiment and Sociality, Advanced Knowledge International,
Adelaide, Australia, pp 19 - 42.
Harvey, I., Di Paolo, E. A., Tuci, E., Wood, R., Quinn, M., (2005). Evolutionary robotics: A new scientific tool for studying cognition. Artificial Life, 11(1/2), pp. 79 - 98.
Iizuka, H. and Di Paolo, E. A. (2008). Extended homeostatic adaptation: Improving
the link between internal and behavioural stability. In From Animats to Animals 10, The Tenth International Conference on the Simulation of Adaptive Behavior, Osaka, Japan, July 7-10, 2008.
|
Dynamics of preference and goal-generation
|
The Ashbyan ideas behind homeostatic adaptation may be combined with an enactive view on autonomy to investigate complex conceptual issues such as spontaneous generation of new goals and the formation of behavioural preferences.
In this view, a preference is not located anywhere in the agent's cognitive architecture, but it is rather a constraining of behaviour which is in
turn shaped by behaviour. Based on this idea, a minimal model of
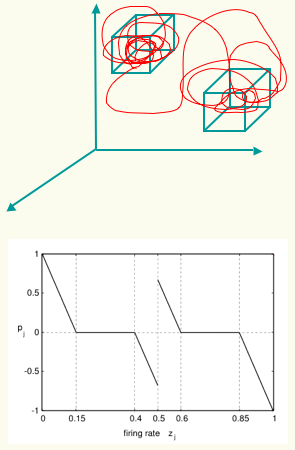
behavioural preference is proposed. A simulated mobile agent is modelled with a plastic neurocontroller, which holds two separate high
dimensional homeostatic boxes in the space of neural dynamics used to regulated synaptic plasticity (right). An evolutionary algorithm is used for creating a link between the boxes
and the performance of two different phototactic behaviours. After evolution, the agent's performance exhibits some important aspects of behavioural preferences such as durability and spontaneous transitions.
These two properties through the mutual constraining of internal and external dynamics in the flow
of alternating high and low susceptibility to environmental variations.
|
|
|
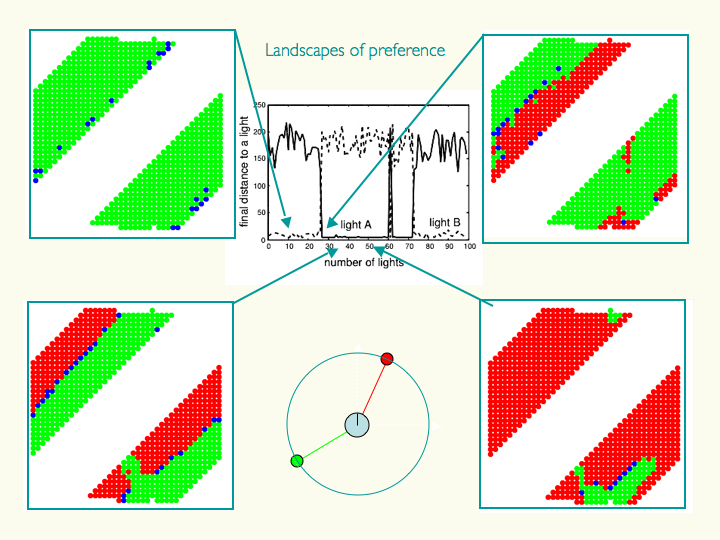
Below: A study of "virtual" choices (what the agent might have done at a given time had the environment been different - in this case, the relative angular orientation of two light sources emitting different coloured light). Plots show the agent's end target of approach depending on the angular orientation of each light (horizontal and vertical axes in each sub-plot) as a function of time over a presentation of 100 pairs of lights. At different points in time the agent shows the alternation between periods of "high commitment" to a goal (one colour is prevalent; agent practically ignores the other option) and periods of environmentally susceptible choice (both choices are highly represented).
Iizuka, H., and Di Paolo, E. A. (2007). Toward Spinozist robotics: Exploring the minimal dynamics of behavioural preference Adaptive Behavior, 15:359-376.
Di Paolo, E. A., and Iizuka, H., (2008). How (not) to model autonomous behaviour BioSystems 91: 409-423, Special issue on Modelling Autonomy.
|
|
Back
|
|